L’ homunculus sensitivo
Il sistema motorio nella visione moderna è strettamente connesso al sistema sensoriale.
Fino ad una ventina di anni fa si aveva la concezione del sistema nervoso come di un sistema con una parte deputata alla percezione degli stimoli esterni col fine di farne dei percetti, la componente sensoriale, che classicamente corrisponde al giro post-centrale dove si colloca l’homunculus sensitivo che rispecchia in modo topografico la densità recettoriale delle diverse parti del corpo e una parte del sistema nervoso che agisce, il sistema motorio, collocato nel giro pre-centrale e a livello del labbro anteriore della porzione della corteccia che è indovata nel solco centrale.
Il cervello associativo del sistema motorio
Nella visione classica queste aree eseguono comandi motori. A cavallo tra questi due “cervelli”, sensitivo e motorio, c’è una terza componente che è detta “cervello associativo”. Nel cervello associativo si includono tutte quelle aree che mettono in connessione e che elaborano ad un livello superiore gli stimoli sensoriali per trasformarli in risposte motorie. In realtà questa visione del cervello distinta in “moduli” è in realtà abbastanza superata poiché negli ultimi venti anni il concetto di sistema motorio si è radicalmente trasformato: oggi si sa che nel sistema motorio non ci sono più semplicemente neuroni che eseguono comandi motori o che danno dei comandi motori ma ci sono neuroni, a livello del s.n.c., hanno funzioni sensori-motorie, vale a dire che nel sistema motorio sono già integrati di per se degli aspetti sensoriali e degli aspetti motori.
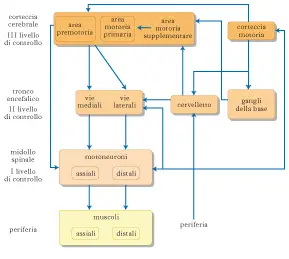
Com’è costituito il sistema motorio
Si diceva che il sistema motorio, ossia quella parte del sistema nervoso che occupa il giro pre-centrale, risultasse costituito da tre aree:
- l’area motoria primaria o area M1
- l’area premotoria
- l’area motoria supplementare
Si parla di aree motorie poiché già nell’Ottocento due studenti di medicina tedeschi avevano scoperto che se si stimola elettricamente la corteccia del giro pre-centrale, si riesce ad ottenere delle risposte motorie nell’emisoma controlaterale dell’animale da esperimento, e rispetto ad altre parti del sistema nervoso l’intensità di corrente necessaria ad ottenere risposta motoria era decisamente minore rispetto ad altre parti del sistema nervoso che pure davano risposte motorie.
La corteccia motoria
In pratica il sistema motorio nasce dall’evidenza che la corteccia che si trova davanti al solco centrale può essere stimolata elettricamente con una minima quantità di corrente per ottenere una risposta motoria che riguardi l’emisoma controlaterale. Da ciò nasce l’idea che esiste una parte del sistema nervoso con funzioni chiaramente ripetitive, il sistema motorio. Successivamente fu dimostrato, sempre alla fine dell’Ottocento, che esiste a livello della corteccia motoria una organizzazione grossomodo topografica, anche se non con il significato moderno, con una specie di mappa delle varie zone che se stimolate facevano contrarre una determinata parte del corpo ma ancora senza la descrizione dell’homunculus motorio di Penfield.
Homunculus motorio di Penfield
Nella prima metà del secolo scorso, questi lavori vengono ripresi da un neurochirurgo canadese, Penfield, nell’uomo, e da un neurofisiologo, Woolsey, nella scimmia. Questi autori dimostrano che a livello del giro pre-centrale c’è una organizzazione somatotopica per cui il corpo risulta rappresentato motoricamente a livello del giro pre-centrale in modo da ottenere una mappa di tutti gli effettori biologici, di tutte le varie parti del corpo, topograficamente e somatotopicamente organizzate. Questa rappresentazione è stata ribattezzata homunculus motorio e prevede che ci sia la rappresentazione della testa a livello del piede del giro pre-centrale e la rappresentazione dell’effettore “bocca” che si spinge fino al giro frontale inferiore; il tronco è rappresentato in una porzione intermedia e i piedi sono rappresentati nella porzione dorsale e occupano anche la superficie mesiale dell’emisfero.
C’è quindi una rappresentazione del corpo sovrapponibile all’homunculus sensitivo del giro post-centrale. Ciò che si sa è che, come l’homunculus sensitivo, l’homunculus motorio ha una rappresentazione dei diversi effettori biologici chiaramente distorta in relazione alla destrezza motorio dell’effettore rappresentato: siccome con le mani siamo in grado di compiere movimenti molto fini questo effettore è rappresentato in modo molto ampio.
Di fatto ciò che è rappresentato è la capacità di un determinato segmento biologico di svolgere dei compiti motori. Allo stesso modo la bocca che ci consente sia di masticare e deglutire ma in particolar modo di articolare suoni per permetterci di comunicare ha una grossa rappresentazione. Il piede che ha una rappresentazione meno ampia e occupa un settore dorsale e un po’ della superficie mesiale dell’emisfero.
Questa rappresentazione è una rappresentazione controlaterale, vale adire che riguarda l’emisoma opposto all’emisfero in esame. Perciò nell’emisfero destro si ritroverà una rappresentazione dell’emisfero di sinistra e viceversa.
La classificazione delle aree fatta da Brodman
Questa rappresentazione somatotopica è presente anche a livello dell’area premotoria, quella che secondo la classificazione di Brodmann è l’area 6, e che si trova al davanti del giro precentrale. La primitiva descrizione somatotopica che è stata fatta a livello dell’area 6 prevedeva che in questa parte del sistema motorio vi fosse una rappresentazione del corpo bilaterale ossia che entrambi gli emisoma avessero una rappresentazione a livello dell’area premotoria. In realtà nell’area 6 i diversi effettori sono rappresentati in modo multiplo e per funzioni diverse.
Questa rappresentazione somatotopica sarebbe presente anche nell’area motoria supplementare e questa area avrebbe un ruolo nella programmazione delle azioni, come pure l’area premotoria, e nella coordinazione delle azioni che richiedono l’uso combinato dei due arti. Sembra che l’area motoria supplementare, cioè quella che è posta a livello della superficie mesiale dell’emisfero, abbia un ruolo specifico nel controllo dei movimenti “bimanuali” e nella programmazione motoria.
Si pensava che le diverse parti del corpo fossero rappresentate a livello di singoli muscoli e che di fatto l’attivazione di queste aree in seguito alla stimolazione dell’area motoria primaria, in seguito alla decisione di muoversi, determinasse il movimento stesso. In realtà per l’area motoria primaria vale l’idea della rappresentazione somatotopica però quello che a livello della M1 è codificato non è tanto l’aspetto esecutivo, che viene deciso in altri ambiti del sistema motorio, ma la forza che deve essere applicata perché si possa eseguire una determinata azione.
Il termine “azione” non è casuale perché si intende con esso una serie di movimenti che vengono eseguiti per raggiungere uno scopo. In effetti tutto ciò che facciamo ha in sé l’obiettivo di raggiungere uno scopo: è raro che si eseguano movimenti non inseriti in atti mirati a raggiungere con uno scopo. Più appropriatamente quindi si deve parlare non di una rappresentazione dei movimenti ma di una rappresentazioni delle azioni.
Un altro aspetto oggi abbondantemente superato è l’idea che il sistema motorio proietti di fatto esclusivamente al midollo spinale e che quindi controlli i movimenti delle varie parti del corpo perché si è scoperto che le maggior parti delle componenti che costituiscono il sistema motorio sono strettamente connesse ad altre parti del sistema nervoso ed in particolare al lobo parietale, più precisamente al lobulo parietale superiore e alle zone intere al solco intraparietale che si trovano nella componente posteriore del lobo parietale, nella costituzione di quelli che oggi sono definiti come circuiti sensori-motori.
I circuiti sensori-motori rispondono a determinate trasformazioni indispensabili per eseguire le azioni.
Quindi nell’idea di sistema motorio classica il sistema motorio esegue dei movimenti con una rappresentazione somatotopica e topografica riguardante sia l’area motoria primaria sia l’area premotoria dove sarebbe presente una rappresentazione bilaterale, e si pensava che nell’area motoria primaria ci fosse una rappresentazione dei diversi movimenti del corpo e che invece l’area premotoria e l’area motoria supplementare avessero un ruolo soprattutto nella programmazione e pianificazione del movimento.
Uno studio di neuro-imaging compiuto all’inizio degli anni ottanta da Roland con la PET, una tecnica che consente di visualizzare le aree che si attivano in relazione a determinati compiti motori e cognitivi sfruttando l’accumulo di sostanze radioattive nei tessuti che svolgono tali funzioni, che dimostrava che in compiti in cui bisognava immaginare di svolgere una determinata azione o programmare una determinata azione si riscontrava una attivazione delle aree mesiali concludendo su questa base che queste aree fossero fondamentali per la programmazione delle azioni.
In realtà oggi si sa che le cose sono un po’ più complicate, infatti si sa che l’area motoria supplementare ha una struttura più complessa tanto che si preferisce definirla come pre-FNA e FNA propriamente detta. Rispetto a questo quadro nelle moderna visuale del sistema motorio sono cambiate alcune cose. Per quanto riguarda l’organizzazione dal punto di vista della suddivisione anatomica, infatti grazie a diverse tecniche citochimiche è stato visto che l’area 6 può essere distinta in diverse sottoaree: c’è una componente ventrale e una componente dorsale.
Le aree del sistema motorio
Nella componente ventrale è possibile poi distinguere due aree inferiori che sono le aree F4 ed F5, dove F sta per frontale e tutte le aree del sistema motorio vengono classificate con questa nomenclatura:F seguita da un numero arabo da 1 a 7. L’area motoria primaria è l’area F1, le altre aree appartengono alla premotoria, o area 6, o alla motoria supplementare. L’area premotoria ventrale può essere distinta nell’area F4 e nell’are F5; l’area F5 in posizione rostrale, l’area F4 in posizione caudale.
Nella porzione dorsale poi si ritrovano l’area F7 in posizione rostrale, e l’area F2 in posizione caudale. In fine sulla superficie mesiale dell’emisfero, ossia nell’area motoria supplementare, si ritrovano l’area F3 e l’area F6.
Oltre alla classificazione proposta, quello che è importante sottolineare è che queste aree hanno delle diverse connessioni con altre parti del cervello: si è visto che molte di queste aree sono connesse con altrettante aree del lobo parietale ed in particolare con alcune aree che si trovano all’interno del solco intraparietale, il quale divide il lobo parietale in lobulo parietale superiore e lobulo parietale inferiore. Nel solco intraparietale si ritrovano diverse aree che risultano connesse in modo stretto con altrettante aree della regione premotoria.
Alcune di queste aree della regione premotoria ed in particolare l’area F6, cioè la pre-FMA (ossia premotoria supplementare mentre invece la F3 è la FMA propriamente detta) e la F7 sono invece legate al lobo prefrontale, la parte del lobo frontale che si trova immediatamente al davanti delle aree motorie.
Mentre invece le altre aree sono connesse con specifiche aree del lobo parietale. Più in particolare l’area F5, quell’area che occupa la porzione rostrale della componente ventrale dell’area premotoria è connessa con un’area che si trova all’interno del solco intraparietale che prende il nome di area AIP (Anterior Intra Parietal).
L’area F4 è legata all’area VIP (Ventral Intra Parietal). Si ritiene che il circuito che unisce la VIP all’area F4 sia un circuito fondamentale per la codifica delle caratteristiche spaziali degli oggetti mentre invece il collegamento tra l’area F5 e l’area AIP sarebbe fondamentale per la codifica delle caratteristiche formali o pragmatiche degli oggetti.
Oltre alla distinzione dell’area 6 nelle varie aree F c’è un’altra area, indicata coma area F8 o FEF (Frontal Eye Field ), che è connessa ad un’area del solco intraparietale chiamata LIP (Lateral Intra Parietal) nella costituzione di un circuito il cui ruolo è quello di codificare i movimenti saccadici, questo sia nel corso della vera esecuzione dei movimenti saccadici, quindi nel porre lo sguardo su un determinato oggetto, sia nella preparazione del movimento, tanto è vero che questa area è ritenuta coinvolta nel paradigma strumentale che prende il nome di “paradigma di Fosner”. Il paradigma di Fosner consiste nel fatto che ci sono dei tempi di reazione facilitati quando si volge lo sguardo in una direzione e si prepara un movimento verso quella direzione.
