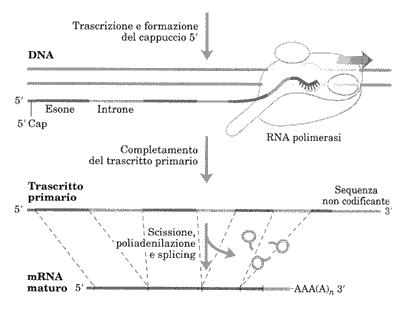
Negli eucarioti c’è una significativa differenza tra geni che codificano per proteine e geni che codificano per RNA. L’elemento promotore più comune nei geni proteici eucariotici è la TATAbox, localizzata tra le posizioni -35 e -20. La sua sequenza consenso TATAAA è molto simile alla regione -10 del sito di aggancio del fattore σ70 procariotico. Un altro elemento del promotore è chiamato iniziatore; la sua sequenza consenso è PyPyAN(T/A)PyPy dove Py indica pirimidina (C o T), N una qualsiasi base e (T/A) significa T o A.
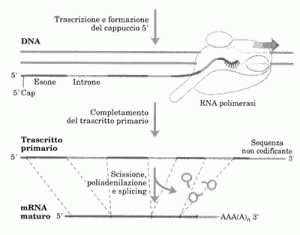
La base A in terza posizione è localizzata in +1, il sito d’inizio della trascrizione. La TATAbox e l’iniziatore sono gli elementi del core del promotore. Ci sono altri elementi spesso localizzati entro 200bp dal sito di trascrizione, come la CAAT box e la GCbox, detti elementi prossimali del promotore.
TATAbox binding protein o TBP
La proteina che interagisce con l’iniziatore e la TATAbox è conosciuta con il nome di TATAbox binding protein o TBP, perché la TATAbox è stata scoperta prima dell’iniziatore. La TBP riconosce non solo il core del promotore di geni proteici, ma anche i promotori di geni per RNA. Questa è una subunità del fattore generale di trascrizione TFIID. Negli eucarioti la trascrizione richiede molti diversi fattori di trascrizione e, nella maggior parte dei casi, fattori di regolazione della trascrizione.
Inizio della trascrizione
Per iniziare la trascrizione la RNA polimerasi eucariotica richiede numerosi fattori di trascrizione che arrivano al sito d’inizio in un ordine specifico. Il TF2D riconosce la TATAbox sul promotore mediante la TBP e vi si lega; il suo legame richiama altri fattori di trascrizione in sequenza quali il TF2A, che si lega sia al promotore che al TF2D, e il TF2B, che si lega al DNA e al TF2A. A questo punto viene richiamate la RNA polimerasiII che lega a sé fattori generali di trascrizione tra cui il TF2F; vengono anche richiamati i fattori 2E e 2H. Tutti i componenti si assemblano sul promotore, interagendo tra loro, con la polimerasi e con il DNA. La doppia elica è aperta dal TF2H, che ha sia attività elicasica che un’altra attività catalitica, infatti è una proteinchinasi capace di trasferire il fosfato in γ dall’ATP a residui di serina sul dominio carbossiterminale della RNA polimerasi, fosforilandola in più siti; ciò induce un cambiamento di conformazione nella coda carbossiterminale della polimerasi che è un segnale per il richiamo di varie proteine che processeranno il m-RNA.
La trascrizione negli eucarioti
I motivi del promotore della RNA polimerasi II sono: l’iniziatore in posizione -2/+4, a monte il nucleo del promotore o TATAbox in posizione -31/-26 e l’elemento BRE di riconoscimento del TFIIB in posizione -37/-32, a valle dell’iniziatore ci possono essere altri elementi come l’elemento del motivo dieci e l’elemento a valle del promotore.
Caratteristiche principali di un enhancer
Le caratteristiche principali di un enhancer sono la distanza, l’orientamento e la posizione, solo la presenza degli isolatori permette di direzionare il segnale; infatti questi ultimi funzionano come marcatori dei confini della cromatina e hanno attività di blocco degli enhancer. Il gruppo di geni che codifica per la β-globina ha, a monte e a valle, due isolatori che impediscono l’espressione di altri geni al di fuori di quelli necessari, l’ehnancer è piuttosto distante dal gene e la regolazione avviene tramite una regione LCR subito a monte dei geni per la β-globina.
Nella regione LCR ci sono siti HS ipersensibili alla Dnasi I che riconoscono fattori di trascrizione specifici, questi fattori si legano tra loro e nelle regioni dell’LCR in maniera coordinata formando un olocomplesso a cui si lega il complesso di preinizio della DNA polimerasi. Questa struttura modula l’espressione del gene e l’espressione dei diversi geni durante la vita tramite le variazioni strutturali e di assemblamento dell’olocomplesso.
Differenti regioni del dna
La trascrizione viene anche regolata da regioni di attacco alla matrice nucleare. Ci sono regioni di DNA che si trovano ancorate alla matrice nucleare (MAR strutturali) mentre altre regioni del DNA hanno la capacità reversibile di ancorarsi alla matrice (MAR funzionali). Quando un TF si aggancia ad un MAR funzionale, questo si ancora alla matrice e induce l’assemblaggio del complesso di preinizio e quindi la trascrizione. Una volta terminata la trascrizione i TF si dissociano e il MAR funzionale si separa dalla matrice.
La RNA polimerasi ha una regione consenso formata da 52 ripetizioni di tirosina, serina, prolina, treonina, serina, prolina, serina (Y, S, P, T, S, P, S). Il complesso proteico mediatore interagisce sia con il complesso di preinizio, con l’istone acetilasi, col complesso di rimodellamento della cromatina ed anche con la proteina attivatrice posta sull’enhancer, determinando un’ansa sul DNA e l’inizio della trascrizione.
Queste proteine attivatrici hanno due domini funzionali:
- uno che riconosce la regione consenso sull’enhancer
- un dominio effettore che riconosce il mediatore e trasmette il segnale.
La proteina attivatrice può avere un dominio di transattivazione, per il legame al mediatore; un dominio di legame al DNA; domini di dimerizzazione, per la formazione di dimeri o tetrameri; un dominio di legame ai ligandi (come il dominio di legame del cAMP nel CAP).
Riassumendo la RNA polimerasi II non contiene una subunità simile al fattore σ dei procarioti, che può riconoscere il promotore e svolgere la doppia elica.
Negli eucarioti queste due funzioni sono svolte da un set di proteine chiamato fattori generali di trascrizione; la RNA polimerasi II è associata a sei fattori di trascrizione (TFIIA, TFIIB, TFIID, TFIIE, TFIIF, TFIIH). Il TFIID è formato dal TBP e dal TAFs, che sono la TATAbox binding protein e i fattori associati; il ruolo della TBP è quello di legarsi al core del promotore e i TAFs aiutano questo processo; nell’uomo i TAFs sono formati da 12 subunità.
Una di queste, la TAF250 ha attività di istone acetiltransferasi, che può rimuovere il legame tra il DNA e gli istoni associati. Il TFIIH è quello con attività elicasica, tuttavia, prima che esso possa aprire il DNA, la RNA polimerasi e almeno cinque fattori di trascrizione devono essere associati a formare il complesso di preinizio.
