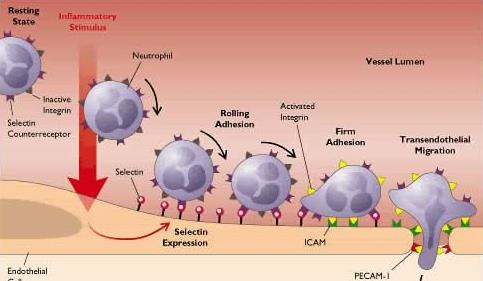
L’uscita delle cellule bianche è un processo attivo e complesso che avviene in stadi successivi e che richiede l’attivazione dell’endotelio in seguito a legame di particolari sostanze.
Nel processo infiammatorio la viscosità del sangue aumenta per sottrazione di plasma. Si osserva che in questa situazione le cellule bianche, che normalmente scorrono al centro della colonna di fluido, tendono a marginare. Quello della marginazione è un processo altamente complesso e selettivo.
L’uscita delle cellule bianche è un processo attivo e complesso che avviene in stadi successivi e che richiede l’attivazione dell’endotelio in seguito a legame di particolari sostanze.
Nel processo infiammatorio la viscosità del sangue aumenta per sottrazione di plasma. Si osserva che in questa situazione le cellule bianche, che normalmente scorrono al centro della colonna di fluido, tendono a marginare. Quello della marginazione è un processo altamente complesso e selettivo.
I globuli bianchi, in particolare i granulociti neutrofili, si spostano in periferia e rotolano lentamente lungo l’epitelio. Questo rotolamento non è un fenomeno prettamente idrodinamico ma prevede l’interazione del leucocita con alcune molecole espresse sulla cellula endoteliale. Lo stadio successivo è quello dell’ “adesione ferma” durante il quale la cellula aderisce tenacemente all’endotelio e comincia a cambiare forma. Infine si passa al fenomeno della diapedesi che consiste nel passaggio attraverso la parete endoteliale: l’endotelio contratto presenta delle piccole finestrazioni che però non sono abbastanza larghe perché possa passare una cellula.
Tuttavia le cellule bianche sono estremamente mobili: la membrana del leucocita comincia ad infilarsi tra due cellule endoteliali e piano piano forzano una separazione maggiore tra di esse in maniera da crearsi un passaggio abbastanza ampio. Nella membrana basale poi cerca delle fenestrature pre-esistenti attraverso le quali possa passare. Infine comincia un “viaggio” lungo anche parecchi mm per raggiungere il sito infiammato.
Un endotelio normale ha pochissima tendenza a legare i leucociti perché il numero di recettori per le molecole adesive dei leucociti (e l’affinità di queste nei leucociti) è molto bassa. Questi recettori sono le P-selectine, normalmente presenti nell’endotelio in granulazioni dette “corpi di Weibel-Palade” ma espresse solo sulle membrane degli endoteli attivati.
Le P-selectine sono molecole di adesione poco efficaci che permettono solo il rotolamento delle cellule. Per cui devono essere sintetizzate (questa volta ex-novo) altre molecole di adesione. Il processo di neosintesi richiede 1-2 ore.
La fase di aderenza stretta dipende:
1. dalla sintesi di molecole di adesione tipo ICAM e VCAM nelle cellule endoteliali attivate;
2. aumento dell’avidità di legame delle integrine (es. LFA1) espresse sulla superficie dei leucociti. Questo processo è stimolato da agenti chemotattici
I leucociti si muovono per chemotassi. Le sostanze chemotattiche possono essere di natura esogena (soprattutto batteriche), prodotte dai leucociti stessi o prodotte da altre cellule endogene.
· Sostanze chemotattiche di origine batterica:c’è un elemento comune a tutti i procarioti ma non agli eucarioti: la formil-metionina, l’aminoacido portato dal tRna che riconosce il codone iniziatore degli mRna procarioti. Le proteine mature vengono modificate post-traduzionalmente ed una delle modificazioni più comuni è il taglio proteolitico del frammento N-terminale, in particolare dei primi 3-4 AA. I leucociti hanno un recettore che riconosce selettivamente piccoli peptidi contenenti formil-metionina all’N-terminale;
· Sostanze chemotattiche di origine endogena:sono prodotte a partire da precursori inattivi presenti nel sangue. Di particolare importanza sono i fattori che si producono dall’attivazione del complemento. Questi fattori devono essere prodotti a partire da C3 o successivi in maniera da coinvolgere sia la via classica che quella alternativa. Infatti i componenti del complemento con maggiore efficacia chemotattica sono C3A e C5A. In particolare C5A è il più potente agente chemotattico essendo infatti attivo anche a concentrazioni di 10-12/-13M;
· Prodotti leucocitari:si tratta di un circuito amplificativo: le stesse cellule che si stanno muovendo in senso chemotattico sono stimolate a rilasciare ulteriori agenti chemotattici. In particolare, altamente potenti sono il leucotriene B4 (anche altri leucotrieni sono chemotattici ma lo sono a concentrazioni molto più alte ed in pratica non hanno significato dal punto di vista fisiologico) e il PAF (o fattore attivante le piastrine. E’ un prodotto del metabolismo fosfolipidico ed ha un’azione molo potente sulle piastrine). La produzione di LTB4 e PAF sono la conseguenza dell’attivazione di altri recettori per agenti chemotattici.
· Chemochine:fino a qualche anno fa questo gruppo era considerato di secondaria importanza. Man mano che la ricerca in questo campo è andata avanti si è però scoperto che le chemochine sono un’ampia famiglia e che svolgono un ruolo importante non solo nell’infiammazione ma anche in molti altri processi (per es. nell’Homing dei linfociti). Le chemochine sono dei peptidi che vengono prodotti da molti tipi di cellule e hanno bersagli diversi. Esse sono raggruppabili in quattro famiglie che hanno in comune dei dettagli strutturali riguardanti l’organizzazione delle cisteine in esse comprese:
a.Famiglia CC:due coppie di cisteine, due cisteine adiacenti, formano due ponti disolfuro per molecola;
b.Famiglia CXC:due coppie di cisteine, due cisteine separate da un aminoacido qualsiasi, formano due ponti disolfuri per molecola;
c.Famiglia C:due sole cisteine e un solo ponte disolfuro per molecola;
d.Famiglia C3XC:due coppie di cisteine, due cisteine separate da tre aminoacidi qualsiasi, formano due ponti disolfuro per molecola.
Si conosce un solo membro di ciascuna delle due ultime famiglie.
Esiste inolte un gruppo di chemochine “costituitive” che presiede alla normale e fisiologica distribuzione dei leucociti nell’organismo; un altro gruppo delle chemochine “inducibili” coinvolte nell’infiammazione e, infine un gruppo di chemochine a cavallo tra i due precedenti gruppi coinvolte nel processo neoplastico.
I recettori per le chemochine sono chiamati semplicemente CCR se il loro ligando è CC e via dicendo.
Più chemochine interagiscono con uno stesso recettore: non vi è assoluta specificità anche se l’affinità di legame sarà diversa.
La sensibilità di una cellula per una chemochina è variabile e dipende dai recettori espressi. Quindi una combinazione di chemochine richiamerà preferenzialmente un tipo di leucociti rispetto ad un altro. Di conseguenza la tempistica di produzione delle diverse citochine nel corso di un processo infiammatorio sarà diversa poiché prima devono interevenire i granulociti neutrofili e, solo in un secondo momento, i monociti e i linfociti.
