Sensibilità nocicettiva
La sensibilità nocicettiva è quella di cui si conosce meno. Si sa che c’è una rappresentazione corticale del dolore, si sa che c’è una componente emotiva che si associa alla sensazione dolorifica, ma non ci sono dettagli circa questa rappresentazione. Dai meccanocettori della cute derivano le informazioni di tipo epicritico che viaggiano attraverso il lemnisco mediale. I recettori superficiali sono quelli di Meissner e Merckel, i recettori profondi sono quelli di Pacini e Ruffini.
Meissner e Pacini sono recettori a rapido adattamento, mentre invece Merckel e Ruffini sono a lento adattamento.
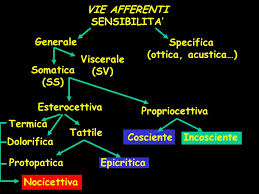
Queste informazioni viaggiano attraverso il lemnisco mediale che si contrappone al lemnisco spinale che invece è la via attraverso cui il fascio spino-talamico porta la sensibilità nocicettiva. Rispetto al decorso classico sono state ultimamente precisate alcune caratteristiche sia delle vie propriocettive sia della sensibilità tattile epicritica: mentre per la via epicritica vale la descrizione classica, cioè di fibre che arrivano al midollo spinale e si portano ai nuclei gracile e cuneato, con un 25% di fibre che invece stazionano a livello delle corna posteriori, per la via propriocettiva c’è una differenziazione tra arti superiori e inferiori in cui le fibre degli arti inferiori non arrivano al nucleo gracile ma alla colonna di Clarck nel midollo spinale e quindi al nucleo Z a livello mesencefalico.
La decussatio lemniscalis
Dal nucleo gracile e dal nucleo cuneato queste fibre si decussano dando luogo alla decussatio lemniscalis che si porta al complesso ventro-basale del talamo. Detto complesso è costituito da due porzioni: la porzione ventro-postero-laterale e la porzione ventro-postero-mediale. Questi neuroni hanno proprietà lemniscali, così come hanno proprietà lemniscali la pars rotunda del nucleo gracile e del nucleo cuneato. Avere proprietà lemniscali significa che i neuroni di queste strutture rispondono ad una solai modalità sensoriale, caratteristica tipica del recettore in quanto tale , hanno dei campi recettivi piccoli, anche se trasducono il segnale di recettori con campi recettivi ampi, e non c’è adattamento, ossia tutte le volte che si determini la stimolazione di questi neuroni essi rispondono.
Rappresentazione somatotopica
A livello del complesso ventro-basale del talamo c’è una rappresentazione di tipo somatotopico: i neuroni sono dedicati a parti specifiche del corpo e in particolar modo lateralmente sono dedicati agli arti inferiori e così via fino ai neuroni mediali dedicati alla faccia. Nel caso della faccia le terminazioni sensitive arrivano dal trigemino. Al complesso ventro-basale del talamo arrivano anche afferenze di tipo nocicettivo. Dal complesso ventro-basale del talamo, i neuroni di terzo ordine proiettano al giro postcentrale della corteccia parietale, il quale risulta costituito di quattro aree: 3a, 3b, 2 e 1.
C’è a questo livello una rappresentazione somatotopicamente organizzata:procedendo dalla porzione mediale verso l’esterno si osserva una organizzazione somatotopica con un homunculus sensitivus deformato, nel senso che la rappresentazione del corpo non è proporzionale alle varie parti del corpo stesso ma è legata alla densità recettoriale di un particolare distretto corporeo. Infatti si riscontra una grossa rappresentazione della faccia e invece una rappresentazione del tronco molto inferiore. Sul versante mediale c’è la zona dedicata all’arto inferiore, mentre invece la componente laterale è dedicata all’arto superiore e alla faccia. All’interno del giro percentuale in relazione alle aree già determinate da Brodmann c’è una suddivisione relata al tipo di sensibilità: le diverse modalità sensoriali sono rappresentate a livello del giro post-centrale in modo segregato, ciò significa che ciascuna modalità sensoriale è rappresentata in modo separato all’interno del giro post-centrale.
Se si considera un taglio del giro postcentrale si osserva che all’interno del solco è disposta l’area 3a e 3 b, sulla convessità l’area 1 e poi al termine della convessità verso il solco postcentrale l’area 2. Questa suddivisione oltre che anatomica è anche di tipo funzionale perché le diverse modalità sensoriali arrivano in modo segregato a queste aree. Più precisamente all’area 3a e 2 arriva a sensibilità di tipo propriocettivo. Alle altre arrivano invece la sensibilità tattile in questo caso con una ulteriore segregazione per quanto riguarda il tipo di recettore, se a rapido o a lento adattamento, quindi corpuscoli di Meissner e Pacini o dischi di Merckel e corpuscoli Ruffini. Anche nel caso del neurone della corteccia del giro post-centrale si riscontrano proprietà lemniscali. A livello della corteccia del giro post-centrale c’è una organizzazione di tipo “colonnare”: se si registra l’attività dei neuroni con elettrodi di registrazione lungo lo spessore della corteccia si ritrovano neuroni incolonnati che rispondono alla stessa modalità sensoriale.
Ciò significa che i neuroni che rispondono ad un certo tipo di sensibilità (quella condotta dai recettori a rapido adattamento piuttosto che a lento adattamento piuttosto che dai propriocettori) è la stessa lungo tutta la colonna di cellule, quindi lungo tutto lo spessore della corteccia cerebrale. La corteccia ha uno spessore variabile da qualche millimetro a 4-5 millimetri. Nel caso del giro post-centrale ci si trova davanti ad una corteccia abbastanza spessa. L’organizzazione colonnare è tipica di tutte le cortecce con modalità sensoriale e per la prima volta è stata scoperta nella corteccia visiva.
Area somatosensoriale secondaria
Oltre all’area somatosensoriale primaria ne esiste una somatosensoriale secondaria che occupa l’opercolo parietale, ossia il piede della circonvoluzione post-centrale. In questa zona c’è una rappresentazione somatotopica per le modalità sensoriali, ma a differenza dell’homunculus sensitivo, che è una rappresentazione essenzialmente controlaterale del corpo, qui si ha una rappresentazione bilaterale. In questa area si ritiene che ci possa essere una integrazione di tipo superiore degli stimoli tattili. Infatti se c’è una lesione di questa area, magari in seguito ad eventi ischemici, quello che si verifica è la cosiddetta “agnosia tattile”, ossia il soggetto non è più in grado di riconoscere gli oggetti attraverso l’esplorazione tattile. Questa area risulta strettamente connessa dal punto di vista anatomico con l’area 6.
Questo non è un caso poiché l’area 6 è un’area al centro di circuiti per l’integrazione senso-motoria: un individuo è in grado di pianificare un programma motorio in relazione alle caratteristiche pragmatiche o alle caratteristiche di posizione spaziale di un determinato oggetto.
Per cui si ritiene che ci possa essere un circuito che dalla area somatica primaria si porta alla area somatica secondaria e dalla somatica secondaria alle aree motorie dedicate alla programmazione motoria. Inoltre l’area somatica secondaria risulta connessa anche con il lobo limbico, e questo fa supporre che questa area somatica secondaria sia responsabile anche in qualche modo degli aspetti emozionali relativi alla stimolazione tattile.
