Il passaggio dei leucociti dall’ambiente ematico nei vasi all’ambiente extracellulare della matrice in cui è presente un focolaio infiammatorio è suddiviso in 4 fasi cronologiche: kemiotassi, marginazione, diaperesi e attacco.
Il passaggio dei leucociti dall’ambiente ematico nei vasi all’ambiente extracellulare della matrice in cui è presente un focolaio infiammatorio è suddiviso in 4 fasi cronologiche: kemiotassi, marginazione, diaperesi e attacco.
1) CHEMIOTASSI
È la attivazione dei leucociti mediante fattori rilasciati da batteri, Interleuchina 1, Complemento, Leucotriene B4, o molecole rilasciati da deperimento cellulare (il tessuto in deperimento è un potentissimo stimolante infiammatorio). È suddivisa in 2 tipi:
chemiotassi temporale: caratteristica delle cellule batteriche prevede una memoria da parte del microrganismo che si muove secondo gradiente chimico del mediatore verso la zona di maggior concentrazione
chemiotassi spaziale: caratteristica dei granulociti che riescono a riconoscere la differenza di concentrazione del mediatore ai capi della membrana (dal davanti alla coda) e si muovono in direzione della concentrazione maggiore.
Il mediatore si associa al recettore di membrana, innescando il rilascio di un II messaggero intracellulare (Ca++) e il conseguente riarrangiamento del citoscheletro emettendo gli pseudopodi necessari a muoversi poi nella matrice.
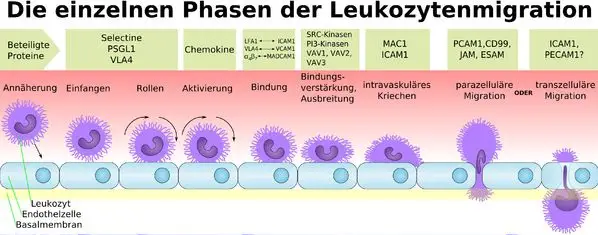
2) MARGINAZIONE
Gli eritrociti si raggruppano in “pile” e occupano lo spazio al centro della colonna del flusso sanguigno nei vasi, facendo spostare i granulociti verso le pareti e quindi a contatto con l’endotelio.
3) DIAPERESI
Dopo la marginazione i leucociti cominciano a rotolare sulle cellule endoteliali, il che comporta la presenza di deboli interazioni tra molecole esposte sulle superfici di entrambe le cellule:
Le cellule endoteliali schierano sulla loro superficie 2 classi di molecole:
SELECTINE P : sono molecole presintetizzate raccolte in vacuoli chiamati Corpuscoli di Weibel-Palade. Quando le cellule endoteliali vengono attivate i vacuoli si fondono con la membrana citoplasmatica e espongono al lume del vaso le selectine.
SELECTINE E : sono molecole che vengono sintetizzate ex-novo quando la cellula endoteliali viene attivata da IL1 e TNF
I Leucociti hanno esposti sulla loro superficie degli zuccheri OLIGOSACCARIDI
Il rotolamento continua fimo a che i leucociti sono completamente adesi alla superficie endoteliale. Ciò comporta ovviamente una diversa interazione, più forte, data da due diversi tipi di molecole:
Le cellule endoteliali espongono le MOLECOLE DI ADESIONE ICAM e DCAM
I Leucociti invece sono dotati di INTEGRINE, molecole che sono sempre esposte sulla loro superficie, ma la cui affinità per le molecole di adesione aumenta quando vengono attivati.
Tutto questo è supportato dall’osservazione che malattie genetiche a carico di geni codificanti integrine o selectine (LAD: Deficienza Adesione Leucociti) portano a problemi di trasferimento dei leucociti dall’ambiente ematico a quello extracellulare, e quindi a gravi infezioni batteriche.
Il fenomeno della diaperesi avviene solo a livello venulare (eccetto nei polmoni in cui la fitta rete capillare lo permette) per non rischiare il blocco del flusso ematico.
L’Istamina induce una retrazione delle cellule endoteliali, facendo aumentare gli spazi interprossimali. I leucociti adesi penetrano in questi spazi e secernendo Elastasi e Collagenasi digeriscono la rete della membrana basale riuscendo a uscire nella matrice extracellulare.
4) ATTACCO
Ha come scopo finale la fagocitosi delle sostanze o dei microrganismi scatenanti il focolaio infiammatorio. La fagocitosi è stimolata da opsonine:
gli anticorpi che sono muniti di 2 siti recettoriali, uno variabile per gli antigeni di superficie batterici e uno costante per antigeni di superficie dei fagociti, si interpongono tra le due specie cellulari mediandone il riconoscimento.
il fattore C3b del complemento che anch’esso riconosce gli antigeni di membrana batterici
le opsonine quindi interagiscono coi granulociti neutrofili e coi macrofagi attivandoli.
Nel fagosoma viene scaricato il contenuto dei granuli primari e secondari che permette la digestione delle sostanze fagocitate.
Può accadere però che il contenuto dei granuli venga riversato nel fagosoma ancora immaturo, quando cioè la membrana citoplasmatica non si è ancora del tutto chiusa sul microrganismo. Allora le molecole rilasciate nello spazio extracellulare diventano mediatori dell’infiammazione, anche danneggiando il tessuto circostante (che diventa necrotico e quindi a sua volta mediatore).
(Fagocitosi di superficie: quando il microrganismo è immobilizzato dal reticolo di fibrina del focolaio infiammatorio)
Con la fagocitosi avviene anche una attivazione metabolica delle cellule fagocitarie, con aumento del metabolismo ossidativo e conseguente consumo di O2 per liberare specie reattive dell’ossigeno tossiche per il microrganismo.
Tutto normale se non fosse che granulociti e macrofagi sono muniti di pochissimi mitocondri e l’energia necessaria al loro metabolismo non deriva da fosforilazione ossidativa ma da glicolisi anaerobia del glicogeno (stivato in granuli nel citoplasma).
Questo è stato appurato somministrando curaro che blocca l’attività mitocondriale. Si è osservato che il leucocita sopravviveva e in fase fagocitaria il metabolismo ossidativo rimaneva invariato. Quindi i mitocondri non intervengono. A cosa serve allora l’O2 che viene consumato?
Nella membrana citoplasmatica delle cellule fagocitarie c’è l’enzima NADPH-ossidasi che in fase di fagocitosi viene a trovarsi anche nelle membrane delle vescicole fagosomiali. Quì utilizza O2 come substrato per formare anione superossido O2- che può avere un effetto dannoso sui lipidi di membrana, e che viene anche dismutato in presenza di H+ in H2O2. Il passaggio da O2- a H2O2 può anche avvenire spontaneamente.
C’è anche l’enzima G6P-deidrogenasi che utilizza G6P in 6-Fosfogluconato dando inizio allo shunt degli esosomonofosfati, innalzando il metabolismo e formando H2O2 da O2.
L’enzima Mieloperossidasi presente nei granuli secondari dei neutrofili, usa come substrati H2O2 + alogeni + H+ per formare Ipoclorito di Na (NaHCl) che è il più potente battericida conosciuto.
In presenza di metalli Cu++ e Fe++ che fungono da catalizzatori, O2- + H2O2 danno un radicale ossidrile OH* che avendo un elettrone spaiato nell’orbitale più esterno è iperreattivo e tende a una forma più stabile. Reagisce così con lipidi di membrana, proteine, DNA, perossidandoli e alterandone le strutture funzionali.
La molecola Glutatione-ridotto trasforma H2O2 in H2O per mezzo di una catalasi.
