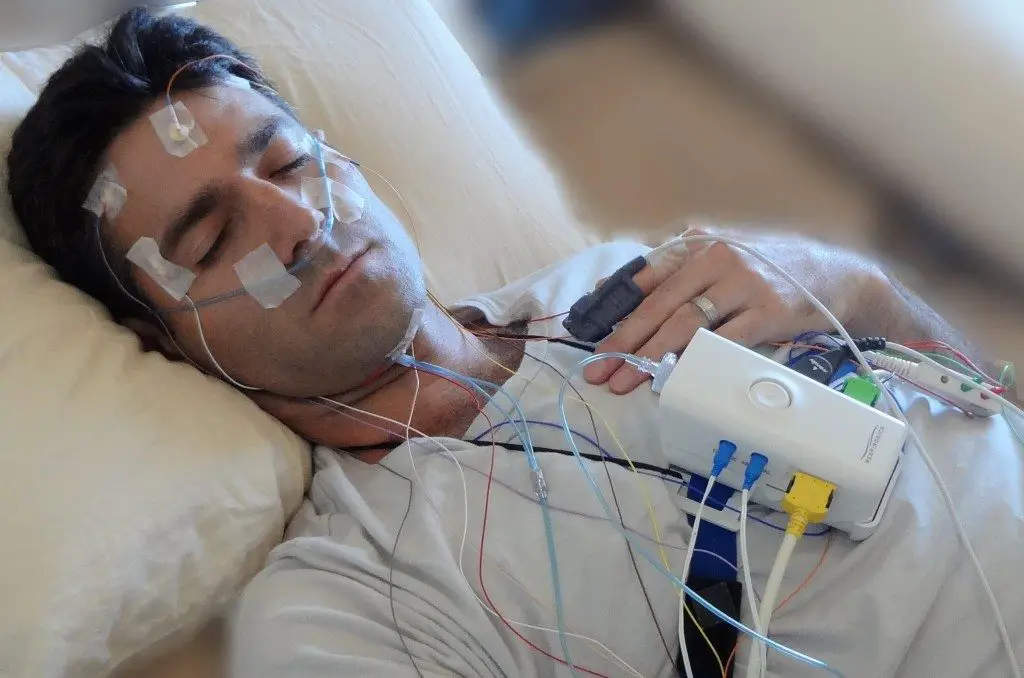
Che cos’è l’Eeg del sonno o polisonnografia
L’EEG serve per studiare il sonno e i suoi disturbi; in questo settore l’EEG prende il nome di “polisonnografia” e questa comprende oltre all’EEG classico, ossia con la distribuzione degli elettrodi secondo il sistema 10-20, anche la registrazione dell’attività muscolare, quindi gli elettrodi di superficie che vengono disposti a livello del muscolo tibiale anteriore, di un elettrodo ai lati degli occhi ed elettrodi diversi a seconda dell’uso clinico. Ci sono tutta una serie di registrazioni che possono accompagnarsi all’EEG di base.

La fase rem e non rem
Sulla base di quello che si registra nell’attività elettroencefalografia, si è visto che il sonno non è uniforme nel corso della notte ma si distinguono due fasi:
- la fase REM
- la fase Non REM
La fase non rem
All’interno della fase non REM si distinguono altri 3 stadi. In uno studio polisonnografico si studia il paziente durante una media di 8 ore di sonno, se invece si vuole focalizzare l’attenzione sulla presenza di attività epilettica nel corso del sonno si restringe la registrazione al primo ciclo di sonno, quindi a due ore di registrazione.
I tre stadi della fase non rem
Il soggetto è invitato in un laboratorio di polisonnografia, si mette a letto per dormire e nei primissimi minuti si avrà una fase di veglia rilassata caratterizzata dalla presenza del ritmo α nelle regioni occipitali, quindi si passa dalla veglia allo stadio 1:
- stadio 1, detto anche stadio di assopimento che è caratterizzato dal fatto che il ritmo α si frammenta e possono subentrare onde un po’ più lente che rientrano nel ritmo θ. In genere l’assopimento dura da 7 a 10 minuti e raramente si ripete nel corso di una notte tipo. Ossia lo stadio uno compare una sola volta nell’arco di una notte tipo perché in genere tutti gli alleggerimenti delle fasi del sonno di una notte tipo, non vanno mai al di sotto dello “stadio 2”, anche quando uno tende a svegliarsi è raro che uno ritorni in stadio 1, a meno che non ci siano dei problemi, come insonnia etc. Quindi lo stadio 1 è caratterizzato da ritmo α rallentato che diventa ritmo θ, dura 7-10 minuti, nell’arco di una notte tipo raramente si ripete.
- La fase successiva è lo stadio 2 che dal punto di vista elettroencefalografico si riconosce per alcuni grafo elementi peculiari: i fusi del sonno, ossia onde molto rapide che si presentano ad una frequenza di 15 cicli al secondo o oltre e con un andamento che è in crescendo/decrescendo, e “i complessi K” che si associano normalmente ad eventi sub-risveglianti nel corso della notte e qualcuno lo definisce anche come “potenziale evocato nel sonno”. Compare nello stadio 2 anche una certa quota di onde δ che però per definizione non può superare il 20% del tracciato, superata questa soglia si entra nello “stadio 3” o sonno ad onde lente.
- Prima, oltre allo stadio 3, c’era anche il 4 e il discriminante era la presenza di onde δ nel tracciato e si diceva che tra il 20 e il 50 % di presenza di queste onde nel tracciato lo stadio era di tipo 3, oltre il 50% si parlava di stadio 4. Una caratteristica dello stadio 3 o sonno ad onde lente è che dal punto di vista di quello che si osserva sul tracciato si ci trova davanti ad un sonno di tipo “sincronizzato”, ossia le onde sono presenti in modo diffuso qualunque sia la derivazione che si consideri.
In tutti gli elettrodi ed in tutte le derivazioni considerate si riscontra un tracciato con presenza di onde δ, per cui sembra che il cervello sia sincronizzato e che le varie parti operino all’unisono. Dopo il sonno profondo si passa allo stadio 2 e quindi si entra in quella fase definita REM, ossia quella fase che nel tracciato si distingue per un taglio netto con il tracciato precedente e un tracciato che assomiglia molto a quello di uno stadio di veglia: “ritmo simil α”, un ritmo α con uno o due cicli in più rispetto al ritmo α basale del soggetto.
La fase Rem
Qualche volta succede che i soggetti non hanno questo rilassamento muscolare e tendono quindi a realizzare l’attività motoria che stanno sognando, questo è un disordine del comportamento di fase REM. Il tracciato quindi si caratterizza per la caduta del tono muscolare, rilevazioni corticali simil-veglia che però si caratterizzano anche per la presenza di alcune onde, definite “onde a dente di sega”, e che sono l’equivalente delle onde ponto-genicolo-occipitale (onde PGO) nell’animale, le quali dipendono da un serie di cellule del tronco dell’encefalo che sono le cosiddette cellule “REM on”, del modello di Mc Cartney ed Obson, che hanno proiezioni verso strutture sovrastanti, quindi corteccia, determinanti la comparsa delle onde PGO, e proiezioni discendenti verso il midollo spinale che determinano la caduta del tono muscolare.
Le PGO originano nel ponte, passano nel corpo genicolato e arrivano a livello del lobo occipitale. Altro elemento che caratterizza dal punto di vista del tracciato il sonno REM sono i movimenti oculari, che danno proprio il nome alla fase. Il sonno REM si associa a tutta una serie di variazioni dal punto di vista autonomico: comparsa di un respiro irregolare, comparsa di aritmie cardiache (tachi o bradicardia), erezione del pene e perdita del controllo termoregolatorio (questa ultima dimostrata solo nell’uomo e consistente prevalentemente nella perdita di termolisi).
L’Ipnogramma
Messo il tutto su un istogramma si ottiene il cosiddetto “ipnogramma”, dove vengono registrati il susseguirsi degli stadi del sonno. C’è un periodo molto breve di veglia, a seguito del quale si riscontra uno stadio 1 che dura da 7 a 10 minuti, quindi uno stadio 2 a cui segue uno stadio 3 (circa 60 minuti), seguito di nuovo dallo stadio 2 e quindi dall’innesco della fase REM. La fase REM compare quindi dopo 70-90 minuti dall’inizio del sonno, questo periodo è importante e prende il nome di “sleep order of REM fase”, cioè soREM e qualcuno aveva ritenuto un soREM più breve del normale un marker biologico di depressione endogena, ma ora questa teoria sembra decaduta.
Gli sleep on set
Addirittura ci sarebbero degli “sleep on set”, ossia l’inizio del sonno ricorrerebbe nella fase REM, ma in realtà questo non è mai stato confermato, semmai è vero che nei “soggetti desincronizzati” ossia privati di elementi esterni che indichino il tempo, è possibile che ci sia la comparsa del sonno nella fase REM. La serie di eventi che va dallo stadio 1 alla fase REM prende il nome di “macrociclo” e il sonno è fatto di 3-4 macrocicli nel corso della notte. Normalmente in un sonno più o meno normale il sonno lento, quindi lo stadio 3, si concentra nei primi due macrocicli mentre negli altri macrocicli prevale il sonno REM, questa è la ragione per cui i sogni del mattino si ricordano di più, ma ciò è vero solo perché in effetti noi sogniamo nella seconda metà della notte.
Il sonno Rem e la temperatura corporea
Alcuni studi hanno dimostrato che il sonno REM va di pari passo con la temperatura corporea, quindi sonno REM e picco minimo di temperatura procedono di pari passo per cui sono probabilmente legati allo stesso tipo di oscillatorie endogeno. Nella prima metà del secolo scorso il sonno era ritenuto un fenomeno passivo e che a determinare il sonno non ci fossero strutture cerebrali specificamente deputate ma che invece siccome durante il corso del buio l’uomo è deprivato di stimoli esterni. In realtà passivo non è per niente, sia perché non è omogeneo nel corso della notte e sia perché ci sono elementi che intervengono in modo attivo atti a controllare tutta una serie di parametri.
C’erano però stati due esperimenti fondamentali compiuti da un fisiologo belga, Bremer, che avevano confermato la tesi che il sonno fosse un fenomeno passivo, almeno secondo una certa interpretazione di questi esperimenti. Bremer aveva studiato l’attività di sonno nel gatto in due preparati animali che erano i cosiddetti “cervello isolato” ed “encefalo isolato”. Nel cervello isolato Bremer faceva una sezione intercollicolare, quindi incideva a livello alto tra strutture encefaliche e tronco dell’encefalo ottenendo la registrazione di una attività cerebrale isolata dalle influenze provenienti dal tronco dell’encefalo. Mentre invece nel preparato dell’encefalo isolato il taglio era tra il tronco dell’encefalo e il midollo spinale ottenendo così il solo isolamento delle informazioni provenienti dal midollo spinale.
La teoria del sonno di Bremer
Bremer misurò l’attività cerebrale con l’elettroencefalogramma e trovò che nel preparato “cervello isolato” l’animale dormiva e il tracciato encefalografico confermava questa condizione, anzi la situazione era pressoché di coma e l’animale era difficilmente risvegliabile. Nel preparato ”encefalo isolato” il tracciato era di veglia, l’animale era reattivo. La conclusione di Bremer era che la ragione per cui nel preparato “cervello isolato” si registra un’attività di sonno consisteva nel fatto che in questo preparato il cervello è isolato da tutte le afferenze sensoriali provenienti anche dai nervi cranici. Quindi se eliminate tutte le afferenze sensoriali l’animale dorme, ciò vuol dire che il sonno è un fenomeno passivo indotto dalla deprivazione sensoriale, tanto è vero che se si lasciano arrivare al cervello le sole afferenze del tronco dell’encefalo, quindi dei nervi cranici, si ottiene che l’animale è sveglio poiché non deprivato dal punto di vista sensoriale.
Gli esperimenti sul sonno di Moruzzi
Per un certo numero di anni questa teoria ha retto e suffragava dal punto di vista sperimentale l’idea che il sonno fosse un fenomeno passivo cioè che fosse legato ad una de-afferenziazione: mi addormento perché di notte sono al buio ed in silenzio. Successivamente ci sono stati tutta una serie si esperimenti compiuti da Moruzzi, un fisiologo italiano, che ha dimostrato invece che la ragione per cui l’animale dorme nel primo caso e rimane sveglio nel secondo è che in realtà nel secondo caso si mantiene l’attività della sostanza reticolare ascendente.
Moruzzi ha dimostrato che in un animale anestetizzato quando si stimola la sostanza reticolare si ha una reazione di risveglio e questo vuol dire che la stimolazione della sostanza reticolare è in grado di determinare una attivazione, quindi un tracciato di veglia, nell’animale, persino quando l’animale è anestetizzato. Moruzzi ha fatto tutta una serie si esperimenti per dimostrare il ruolo attivatorio della sostanza reticolare ascendente ma in particolare c’è stato un preparato, il preparato “mediopontino-pretrigeminale”, in cui, siccome si salva il trigemino, ci si aspetterebbe , secondo l’ipotesi di Bremer, che l’animale rimanga vigile e invece l’animale dorme.
L’animale dorme perché non c’è più l’influenza attivatrice della sostanza reticolare ascendente che rimane in un livello inferiore al taglio che non può quindi indurre attivazione sui nuclei aspecifici del talamo e quindi in modo diffuso sulla corteccia. Questa è la dimostrazione che il sonno non è un fenomeno passivo, ma in realtà esistono nel cervello delle strutture che specificamente sono deputate a mantenere la veglia e a determinare se inibite il sonno.
Studi successivi hanno dimostrato che all’interno della sostanza reticolare c’è una porzione ascendente, descritta da Moruzzi, in grado di determinare la veglia e una parte caudale, costituita da diversi nuclei, che invece avrebbe una funzione specificamente “ipnoinducente”. Il passo ulteriore che si è compiuto oggi è quello che si considera il sonno e la veglia come due aspetti di un unico sistema che è il sistema che garantisce il “sistema di vigilanza” dell’individuo che ha un andamento circadiano ed è evidentemente influenzato da “indicatori di tempo” esterni, in particolare la luce.
Come tutti i ritmi circadiani hanno un’organizzazione per cui va considerato un elemento di sincronizzazione che è chiamato “indicatore di tempo”. C’è un recettore a livello retinico, distinto dai fatocettori responsabili della visione, che da’ origine ad una via di sincronizzazione chiamata “fascio retino-ipotalamico” il quale arriva al nucleo soprachiasmatico dell’ipotalamo il quale è di fatto l’orologio biologico più importante dell’organismo e dal quale dipendono tutta una serie di altri orologi, non bene definiti in realtà, che probabilmente controllano le diverse funzioni.
Quindi avremo, rispetto al pace-maker che è il nucleo soprachiasmatico dell’ipotalamo tutta una serie di “oscillatori schiavi” che lavorano in relazione a quanto stabilito dal nucleo soprachiasmatico. Il nucleo soprachiasmatico dell’ipotalamo è legato alla epifisi che produce melatonina, l’ormone il cui picco viene registrato nella fase che precede il sonno e che probabilmente lo induce, tanto è vero che è ormai di moda utilizzare la melatonina come fattore di sincronizzazione per quelle persone che viaggiano molto e sono soggette a diversi fusi orari e quindi a sbalzi di abitudini che inducono una difficoltà a riprendere il ciclo sonno-veglia.
