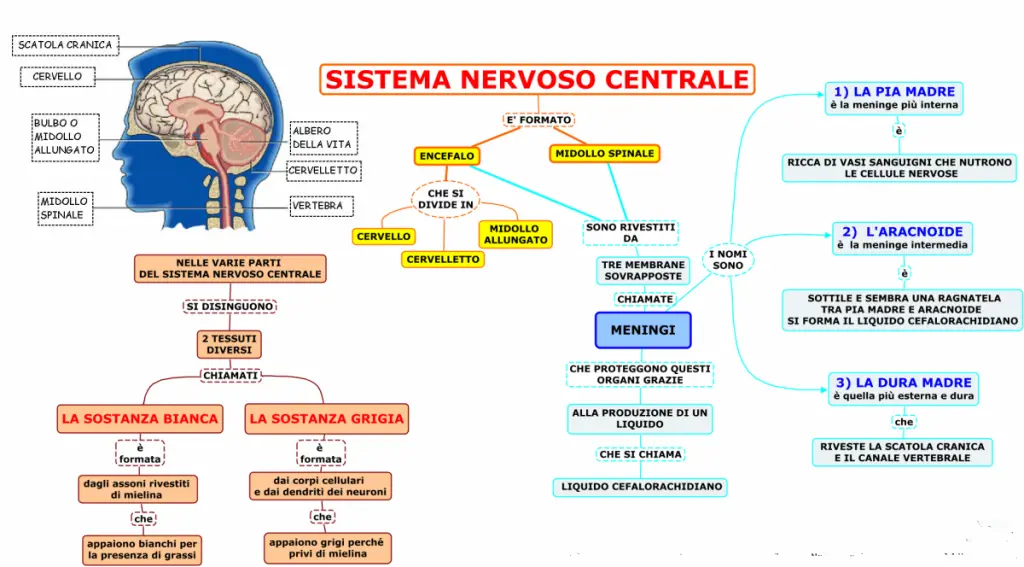
Il sistema nervoso è costituito da neuroni e le cellule gliali, con svariate funzioni. Glia significa “colla” e le cellule gliali tradizionalmente sono stati considerati elementi di sostegno.
Gli elementi gliali sono 5 e ognuno di essi ha una o più particolarità funzionali:
• Microglia ha funzione fagocitaria;
• Astrociti formano la barriera emato-encefalica e hanno un ruolo fondamentale nel controllo della stabilità dei liquidi extracellulari, più nello specifico gli astrociti si dispongono in modo da “tappare” con i loro prolungamenti citoplasmatici i pori tra le cellule endoteliali del vaso, di modo da costituire un filtro per le sostanze presenti nel circolo ematico;
• Cellule ependimali, rivestono il lume dei ventricoli cerebrali e provvedono alla produzione del liquido cefalorachidiano;
• Oligodendrociti producono la mielina del sistema nervoso centrale, che si dispone a rivestire una porzione di più assoni da un nodo di Ranvier al successivo di più neuroni;
• Cellule di Schwann svolgono la stessa funzione degli oligodentrociti ma a livello del sistema nervoso periferico avvolgendo un singolo assone.
La mielina non è altro che il prolungamento della membrana citoplasmatica che si avvolge intorno all’assone, ha una funzione fondamentale nel sistema nervoso poiché funge da isolante e permette all’impulso di saltare da un nodo di Ranvier, ossia una zona di contiguità tra due mieline isolanti lo stesso assone a contatto diretto con il liquido extracellulare, aumentando in tal modo la velocità di conduzione dell’impulso nervoso. Ecco perché le fibre mieliniche hanno un tipo di conduzione, detta di tipo saltatorio, più veloce delle fibre amieliniche in cui la conduzione è di tipo “elettrotonica”.
A livello dei nodi di Ranvier c’è un’alta concentrazione di “canali voltaggio-dipendente” per il Na+ e per il potassio che fanno in modo che la trasmissione dell’impulso proceda da un nodo all’altro. Inoltre la mielina aumenta il diametro della fibra nervosa e questo è un ulteriore fattore che facilita la trasmissione dell’impulso.
I neuroni sono costituiti da:
• Soma, o pirenoforo;
• L’assone, il prolungamento che trasmette in senso centrifugo l’informazione o l’impulso anche a metri di distanza;
• I Dendriti rappresentano la componente del neurone che riceve l’informazione o l’impulso e non si allontana molto dal soma, ma ha un sistema di arborizzazioni molto estese.
Dal punto di vista strutturale i neuroni possono essere distinti in:
Unipolari;
Pseudo-unipolari, con neurite a T;
Bipolari;
Multipolari.
Dal punto di vista funzionale i neuroni si distinguono in:
Motoneuroni, o efferenti;
Neuroni sensitivi o afferenti;
Interneuroni, o neuroni di connessione.
Nei neuroni, come in tutte le altre cellule dell’organismo, esiste una differenza di potenziale tra l’interno della cellula e il suo esterno: normalmente l’interno della cellula è più negativo rispetto all’esterno. . Questa differenza di potenziale è definita come “potenziale di riposo” della cellula e nei neuroni è pari a -70mV con l’interno più negativo dell’esterno. Il potenziale di riposo è la differenza di potenziale che si instaura fra interno ed esterno della cellula.
Per le cellule come i neuroni e le cellule muscolari è bene usare la terminologia di ”potenziale di riposo” poiché esse possono variare la differenza di potenziale ai lati della membrana; sono dette cellule “eccitabili”, ossia in grado, sotto opportuni stimoli, di modificare il proprio potenziale di membrana. Questa modifica si può manifestare come “potenziale graduato” o come “potenziale d’azione “ ed è la modalità con cui queste cellule comunicano tra loro. Il potenziale di riposo si può modificare o aumentando la differenza di negatività interna rispetto all’esterno della cellula, si parla in questo caso di “iperpolarizzazione”, oppure diminuendola, si parla in questo caso di “depolarizzazione”.
Poiché la cellula nervosa per poter comunicare con altre cellule si deve depolarizzare, tutti gli stimoli che di fatto rendono l’interno della cellula meno negativo, fanno si che la cellula proceda verso un potenziale d’azione, tutti gli stimoli che rendono più negativo l’interno della cellula, fanno in modo che la cellula si allontani da una condizione di eccitabilità propria del potenziale d’azione.
Il potenziale di riposo di una cellula nervosa è di -70 mV, e ciò è posto in essere da tre fattori principali:
1) Una diversa distribuzione di cariche fra esterno e interno della cellula, legata alla diversa distribuzione delle concentrazioni degli ioni Na+, K+ e Cl-;
2) Una diversa permeabilità della membrana ai suddetti ioni, in merito a diversi canali. Tali canali sono distinguibili come:
a. Canali passivi, sempre aperti, che lasciano passare gli ioni in modo libero, seguendo il gradiente di concentrazione dello ione, sono più permeabili allo ione K+ che non allo ione Na+ con un rapporto di 25:1;
b. Canali ligando-dipendenti, che modificano la propria conformazione, quindi, la propria permeabilità agli ioni, quando si uniscono a specifiche sostanze, i neurotrasmettitori;
c. Canali voltaggio-dipendenti, che modificano la propria permeabilità ad uno specifico ione in relazione ad una variazione del potenziale di membrana.
3) Un sistema di pompe attive che fa si che il gradiente ionico tra esterno ed interno si mantenga costante. La pompa principe è la Na+/K+ ATPasi dipendente essa mantiene costante la concentrazione degli ioni sodio e potassio ai lati della membrana. E’ un sistema che va contro gradiente e pertanto implica un dispendio di energia, in questo caso l’energia utilizzata è ricavata dalla idrolisi dell’ATP. La pompa è in grado di legare 3 ioni Na+ dal liquido intracellulare e ciò facilita l’idrolisi dell’ATP e in queste condizioni avviene un cambiamento conformazionale rivolgendovi verso l’esterno dove lascia gli ioni Na+ e lega 2 ioni K extracellulari, rilasciando così il gruppo fosfato, e ricambiando conformazione, portando gli ioni K all’interno della cellula.
A scopo didattico si assume che la cellula sia permeabile solo ad uno di questi ioni, il Na+. Lo ione Na+ risulta più concentrato all’esterno della cellula rispetto all’interno. In queste condizioni segue il gradiente di concentrazione, chiamato anche ”forza chimica”, e lo ione Na+ tende ad entrare all’interno della cellula, che deve essere permeabile solo a questo ione.
Poiché lo ione è per definizione una particella con carica, in questo caso positiva, si viene a creare una condizione in cui la “forza elettrica”, venutasi a creare all’interno della cellula dove ci sono solo ioni carichi positivamente, tenderà a respingere il flusso di cariche positive diretto all’interno della cellula.
Nel tempo si verrà a creare un equilibrio dinamico tra ioni che seguono il gradiente di concentrazione e quindi entrano nella cellula e particelle che seguono il gradiente elettrico e quindi escono dalla cellula. In questa situazione c’è un flusso netto di cariche nullo e si dice che quello è il “potenziale di equilibrio per quello specifico ione “. Il potenziale di equilibrio per lo ione Na+, se si assume questo come unico ione a cui la cellula è permeabile sarebbe di +60 mV. L’ equazione di Nernst, esprime il potenziale di equilibrio di uno specifico ione, se di questo ione sono note la sua carica e la sua concentrazione ai lati della membrana.
Iκ= RT log [I]e
ZF [I]i
Dove:
Iκ= potenziale di equilibrio ione,
R= costante dei gas,
T= temperatura in Kelvin,
Z= la valenza dello ione,
F= la costante di Faraday.
Il potenziale di equilibrio di uno ione dipende dalla sua carica e dalla diversa concentrazione che lo ione ha all’interno e all’esterno della cellula, nella condizione in cui la membrana sia selettivamente permeabile solo per questo dato ione. Nel caso lo ione sia il K+, è più concentrato all’interno della cellula, si avrà una fuoriuscita che ad un certo punto verrà tamponata dal gradiente elettrico e ci sarà un passaggio netto di cariche nullo, quindi, lo stato di potenziale di equilibrio del K+ che sarebbe uguale a -94 mV.
Nella realtà la cellula è permeabile a più ioni e il reale potenziale di riposo della cellula è -70 mV, quindi, più vicino al potenziale di equilibrio dello ione K+ rispetto al potenziale di equilibrio dello ione sodio.
Ciò si spiega:
I canali passivi per gli ioni sono più permeabili allo ione K+ che allo ione Na+: di fatto il potenziale di riposo dipende dallo ione K+ più di quanto dipende dallo ione Na+;
L’equazione di Hodgkin-Goldman-Katz corregge il potenziale di equilibrio calcolato con l’equazione di Nernst tenendo conto della permeabilità diversa della membrana diversa per gli ioni Na+ e K+ e della concentrazione degli ioni all’interno e all’esterno della cellula.
RT Pk [K+]e + PNa [Na+]e + PCl [Cl-]i
Vm = —— ln ——————————————————
F Pk [K+]i + PNa [Na+]i + PCl [Cl-]e
Il potenziale di riposo può essere modificato dall’applicazione di uno stimolo, che può provenire da una struttura preposta a recepire informazioni dal mondo esterno o da un’altra cellula. La risposta della cellula può essere una iperpolarizzazione o una depolarizzazione. Il potenziale graduato è una modifica del potenziale di riposo di una cellula nervosa, nel senso di una iperpolarizzazione o di una depolarizzazione, in entrambi i casi, però, non sufficiente a generare un potenziale d’azione.
Inoltre i potenziali graduati:
• Possono sommarsi, in modo algebrico, tra di loro per raggiungere il valore soglia, oltre il quale si ha la genesi del potenziale d’azione. Questa somma algebrica può riguardare variazioni del potenziale di riposo in senso iperpolarizzante e in senso depolarizzante. Inoltre, questa sommatoria può essere in senso spaziale, cioè si possono sommare due potenziali d’azione insorti in punti differenti della cellula; in senso temporale, cioè due potenziali graduati che si avvicendano nel tempo.
• I potenziali graduati sono dipendenti dall’intensità detto stimolo: un aumento dello stimolo fa aumentare il potenziale graduato;
• I potenziali graduati posso generarsi in punti diversi dal monticolo assonale;
• I potenziali graduati non seguono la logica del tutto o nulla, ma sono legati all’intensità dello stimolo;
• Il potenziale graduato si risolve localmente, cioè non si trasmette;
• Esempi di potenziali graduati sono potenziali post-sinaptici di tipo eccitatorio o inibitorio o potenziali di trasduzione, fino a quando almeno non raggiungono una certa soglia.
Il potenziale d’azione è un evento stereotipato, cioè che si ripete costante nel tempo, che si genera seguendo la legge del tutto o nulla, inoltre è in grado di diffondere attraverso l’assone del neurone, sino all’elemento cellulare successivo:
• Insorge solo e esclusivamente nel monticolo assonale, struttura che funge da centro di integrazione per i vari potenziali graduati sino al raggiungimento del valore soglia che porta al potenziale d’azione stesso;
Nel potenziale d’azione si ha una modifica del potenziale di riposo che si verifica in 3 momenti distinti:
a) Depolarizzazione rapida, di 1 ms;
b) Ripolarizzazione rapida, di 1 ms;
c) Iperpolarizzazione postuma, che può protarsi sino a 15 ms.
Nella fase di depolarizzazione rapida il potenziale passa da -70mV fino a +30mV, quindi l’interno della cellula diventa positivo rispetto all’esterno, tutto ciò in 1ms.
Nella ripolarizzazione rapida si ritorna a -70mV.
Nella iperpolarizzazione postuma si scende ad un valore ancora più negativo, stato che si protrae per un periodo di tempo che può arrivare a 15ms.
Nel corso del potenziale d’azione si verifica una “modifica della permeabilità di alcuni canali voltaggio dipendenti” per gli ioni Na+ e K+.
Il Na+ essendo più concentrato all’esterno tende addentrare nella cellula e il K+ viceversa tende ad uscire. E’ stato formulato un modello per spiegare il funzionamento dei canali voltaggio dipendenti per lo ione sodio e per lo ione potassio.
Il meccanismo di un canale voltaggio-dipendente per il Na+ sarebbe legato alla presenza all’interno di questo canale di due cancelli, e cioè un cancello di attivazione e un cancello di inattivazione. In condizione di riposo, ossia ad un voltaggio di -70mV, in cancello di inattivazione è aperto e in cancello di attivazione è chiuso. Quando l’integrazione dei vari potenziali graduati ha fatto generare il potenziale d’azione, in cancello di attivazione si apre e il passaggio di ioni Na+ all’interno della cellula giustifica il passaggio da -70mV a +30mV .
Nel secondo ms la curva ridiscende nel momento in cui si richiude la porta di inattivazione (fase di ripolarizzazione rapida ), cioè quella che in condizioni di riposo è aperta, in questo modo il Na+ non può più fluire all’interno della cellula. Allo stesso tempo si aprono i canali voltaggio-dipendenti per il K+, per cui si ammette esista una sola porta, che si attiva nella fase di ripolarizzazione, e che continua ad essere aperta anche quando la cellula ha raggiunto di nuovo il valore del potenziale di riposo a -70mV. Questa persistenza di apertura della porta del canale voltaggio-dipendente del canale del K+ giustifica la iperpolarizzazione postuma, ossia la discesa a valori inferiori a -70mV dovuta all’eccessiva fuoriuscita di cariche positive dalla cellula.
Nel potenziale d’azione non c’è differenza tra uno stimolo più o meno intenso poiché, dal punto di vista della genesi, basta raggiungere il potenziale soglia per avere una risposta stereotipata. Per riuscire a capire se uno stimolo è più o meno forte di un altro, se è uno stimolo soglia, soprasoglia, uno stimolo forte, uno stimolo fortissimo, il sistema nervoso fa riferimento al “codice di frequenza”. Ossia uno stimolo sopra soglia è in grado di determinare un numero, quindi una frequenza, maggiore di potenziali d’azione, rispetto a uno stimolo soglia, ed è il numero di potenziale d’azione che discrimina un certo stimolo più forte di un altro.
Per le sue caratteristiche l’evento “potenziale d’azione“ non può ripetersi nel tempo in modo invariato, cioè esiste un periodo nel quale se si è stabilito un potenziale d’azione, non se ne può generare un altro, questo periodo è il cosiddetto “periodo refrattario assoluto”. Il periodo refrattario assoluto cade in quei circa 2 ms che vanno dalla depolarizzazione rapida alla iperpolarizzazione rapida, e corrisponde al periodo in cui è impossibile per la cellula generare un potenziale d’azione. Ciò si giustifica pensando allo stato dei canali voltaggio dipendenti nelle fasi appunto di depolarizzazione rapida e iperpolarizzazione rapida, che in questa fase sono già massimamente attivati e le porte sono già tutte aperte.
C’è però il periodo che coincide con l’ultima fase di iperpolarizzazione rapida e la fase di iperpolarizzazione postuma in cui è possibile, se si somministra uno stimolo sopra soglia, generare un nuovo potenziale d’azione, poiché in questa fase i canali del Na+ sono chiusi e possono essere riaperti dallo stimolo sopra soglia. Il periodo refrattario relativo si associa alla possibilità di generare nuovi potenziali di azione, e dal numero, ossia dalla frequenza dei potenziali d’azione generati il sistema nervoso è in grado di comprendere l’intensità dello stimolo.
Il potenziale d’azione, una volta generato, si propaga con effetto domino lungo tutto l’assone. La modalità di propagazione è diversa a secondo che l’ assone sia di tipo mielinico o amielinico. Poiché nel primo caso la conduzione è di tipo “saltatorio”, e poiché i canali voltaggio-dipendenti sono concentrati in prossimità dei nodi di Ranvier, nelle fibre amieliniche la conduzione è di tipo “elettrotonico”. Quando si ha un punto della membrana in cui si è generato un potenziale d’azione o un punto della membrana in cui c’è un’inversione della polarità della cellula, questo evento fa si che ci sia un richiamo di cariche positive dall’esterno, dove si è creato un polo negativo e viceversa un passaggio di cariche negative verso la zona all’interno della membrana cellulare che è diventata positiva.
Questo fa si che l’inversione della polarità della membrana proceda lungo tutto l’assone fino al neurone successivo, essa è detta conduzione elettrotonica ed avviene nelle fibre amieliniche. La ragione per la quale la conduzione si propaga soltanto in una direzione è dovuta al periodo refrattario assoluto del potenziale d’azione, quindi, lo stimolo può solo proseguire nella direzione in cui ci sia una membrana che non sia nello stato di refrattarietà, ossia verso il neurone successivo. Quindi questa conduzione è detta “unidirezionale” e assomiglia all’effetto domino.
Diversa è la situazione per le fibre mieliniche. In questo caso i canali voltaggio-dipendenti si concentrano in prossimità dei nodi di Ranvier, questo fa si che la conduzione sia di tipo saltatorio. In questo caso la velocità di conduzione è più veloce sia per la conduzione di tipo saltatorio sia perché il diametro della fibra maggiore e quindi oppone una minore resistenza.
